Aaron Chevalier # 1 2, Tao Guo # 1 3, Natasha Q Gurevich # 1 2, Jingwen Xu 1 3, Masanao Yajima 3, Joshua D Campbell 1 2
Affiliations Expand
- PMID: 40778578
- PMCID: PMC12395221
- DOI: 10.1158/2767-9764.CRC-24-0496
Abstract
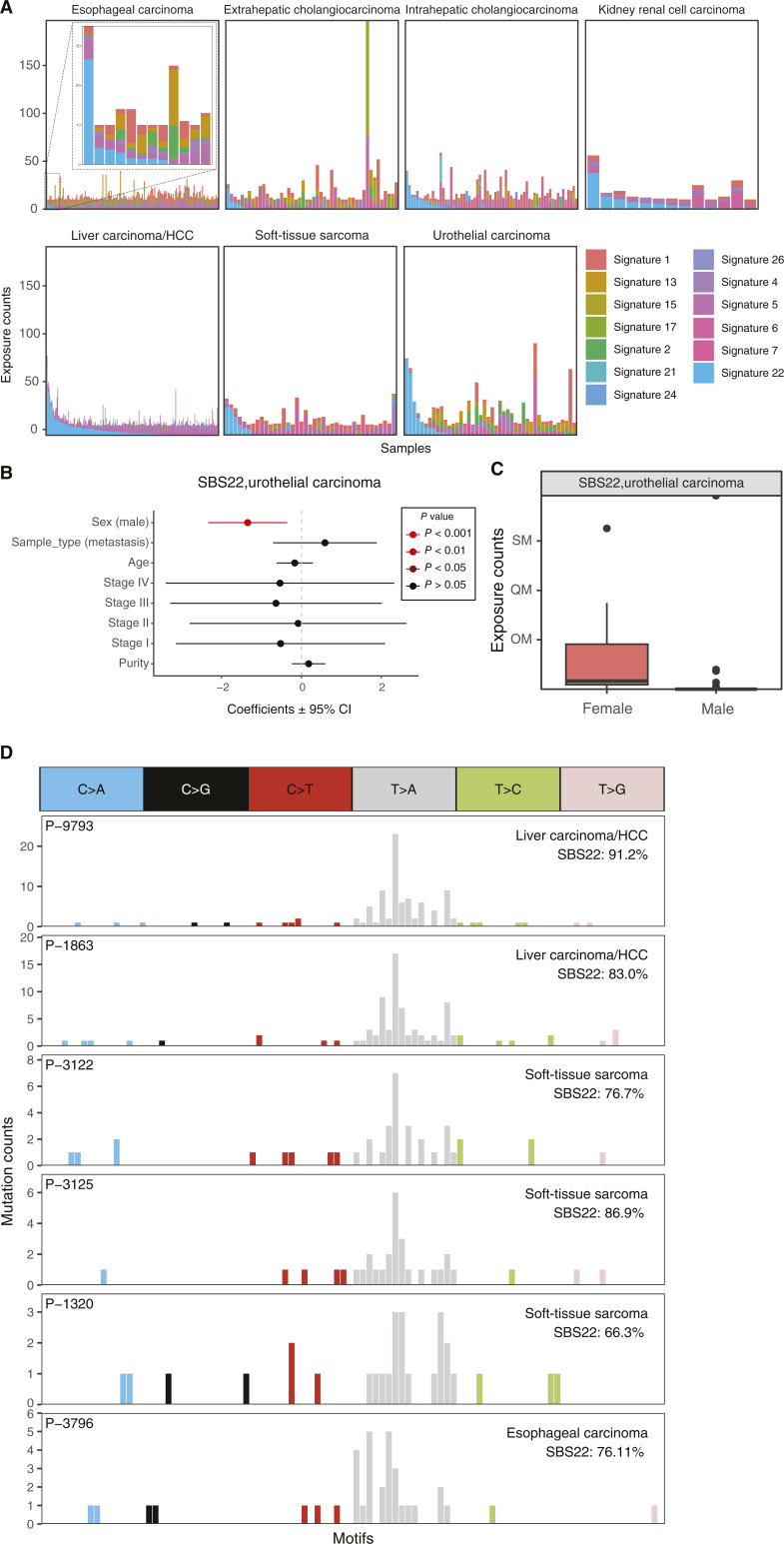
The majority of mutational signatures have been characterized in tumors from American and European countries, and the degree to which mutational signatures are similar to or different from those in Chinese populations has not been fully explored. We leveraged a large-scale clinical sequencing cohort of tumors from the Chinese population developed by OrigiMed (OM). A total of 2,115 tumors from 25 major tumor types that had at least 10 single-base substitutions (SBS) were used for mutational signature analysis. Sixteen mutational signatures from the Catalogue of Somatic Mutations in Cancer (COSMIC) database were identified in the OM cohort using the musicatk package. The activity levels for most of the signatures were similar to those in a cohort from Memorial Sloan Kettering in the United States. The activity level for the apolipoprotein B mRNA editing catalytic polypeptide-like-related signature SBS2 was significantly lower in breast tumors in the OM cohort. The aristolochic acid signature SBS22 was observed in four soft-tissue sarcomas, and the POLE-associated signature SBS10 was observed in a gallbladder carcinoma. In lung adenocarcinoma, the polycyclic aromatic hydrocarbon signature SBS4 was significantly higher in males compared with females but not associated with smoking status. UV-related mutations were significantly lower in cutaneous melanomas from the Chinese cohort compared with the American cohort and may contribute to lower response rates to immunotherapy observed in this population. Overall, these results add to our understanding of the mutational processes that contribute to tumors from the Chinese population.
Significance: Analysis of a large Chinese cohort from 25 tumor types reveals similarities and differences in the activity of mutational signatures compared with other populations.
©2025 The Authors; Published by the American Association for Cancer Research.
Conflict of interest statement
No disclosures were reported.
Figures